Dirk’s et al proposed in 2015 that the remains had were deliberately placed in the chamber in an act of deliberate and repeated disposal of the dead by other naledi individuals. While this hypothesis was seen as controversial by a few researchers it has yet to be disproved and remains the presently supported theory on the way in which naledi individuals found their way into the chambers and passages.
In 2017 it was reported that initial work on dating sediments and geological formations within the Dinaledi subsystem, as well as direct dating of individual naledi specimens resulted in the proposed dates of 335,000 and 236,000 years ago. It is important to note that this age range is only an estimate of the fossil remains in the Dinaledi and Hill Antechambers and does not reflect the presently unknown whole temporal range of the species.

Homo naledi brain anatomy, however, is somewhat like contemporary Homo, which may allude tocognitive complexity in the species. However, in the absence of direct association of tools, while many ascribe the likelihood that Homo naledi should be associated with contemporary Middle Stone Age lithic industries, others disagree based largely on the perception that brain size is directly associated with cognitive abilities and the development of more advanced lithic technologies. However, the persistence of a small-brained hominin in Africa during what was previously thought to be a temporal period when only bigger-brained hominins existed, at the very least revises previous ideas that a larger brain was an inevitable consequence of later Pleistocene hominin evolution.
Homo naledi’s postcranial anatomy indicates that they were capable and competent at long-distance walking possessing a humanlike stride and gait, although there are differences in their pelvic and hip anatomy from humans. Naledi was clearly also however, a capable climber and better adapted than humans to climbing and suspensory behaviours. Their hands demonstrate long, opposable thumbs and curved fingers.
Their dental anatomy is generally primitive and in fact unique, and surface wear studies suggests consumption of gritty and coarse foods showing similar abrasion and pitting to that seen in robust australopithecines, although with teeth closer to Homo sapiens in size.
Although charcoal had been noted previously as being present in the Lesedi Chamber of the Rising Star system, it was not associated directly with Homo naledi at the time of its description. Berger and his colleagues now report widespread evidence of controlled use of fire for lighting and the cooking of food throughout the whole of the Rising Star cave system and associate this evidence of hearths, charcoal, ash and burnt bones to Homo naledi being capable of making and transporting fire indicating a higher level of both cognitive ability than some have attributed to the species and also offering an explanation for how Homo naledi was accessing such remote subterranean spaces safely.
Skull
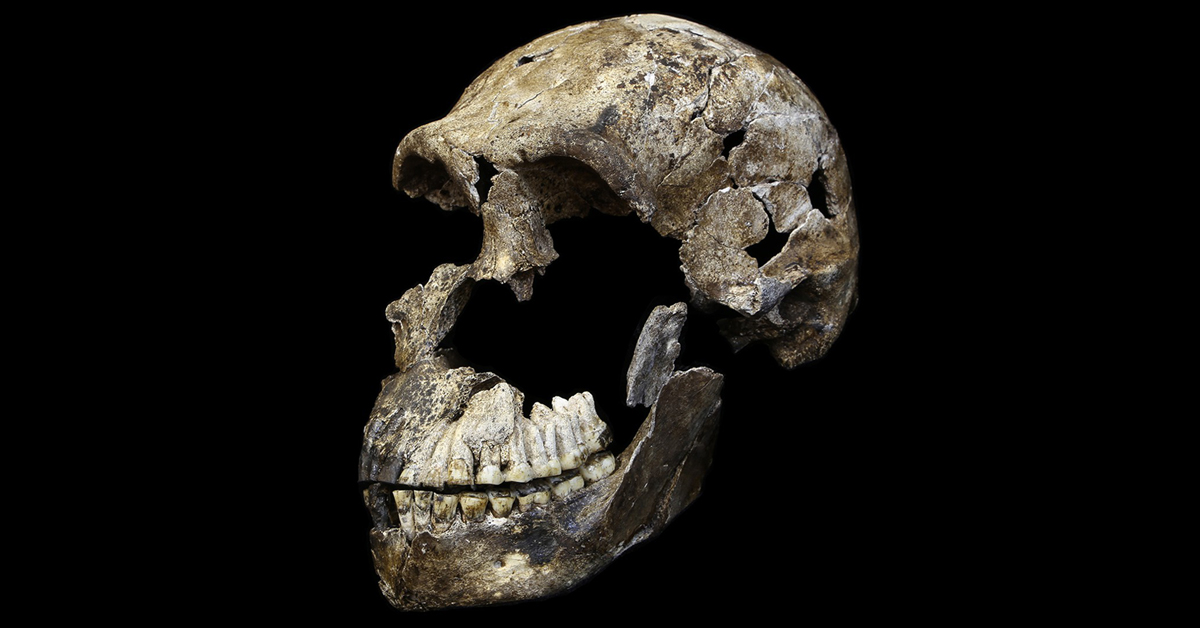
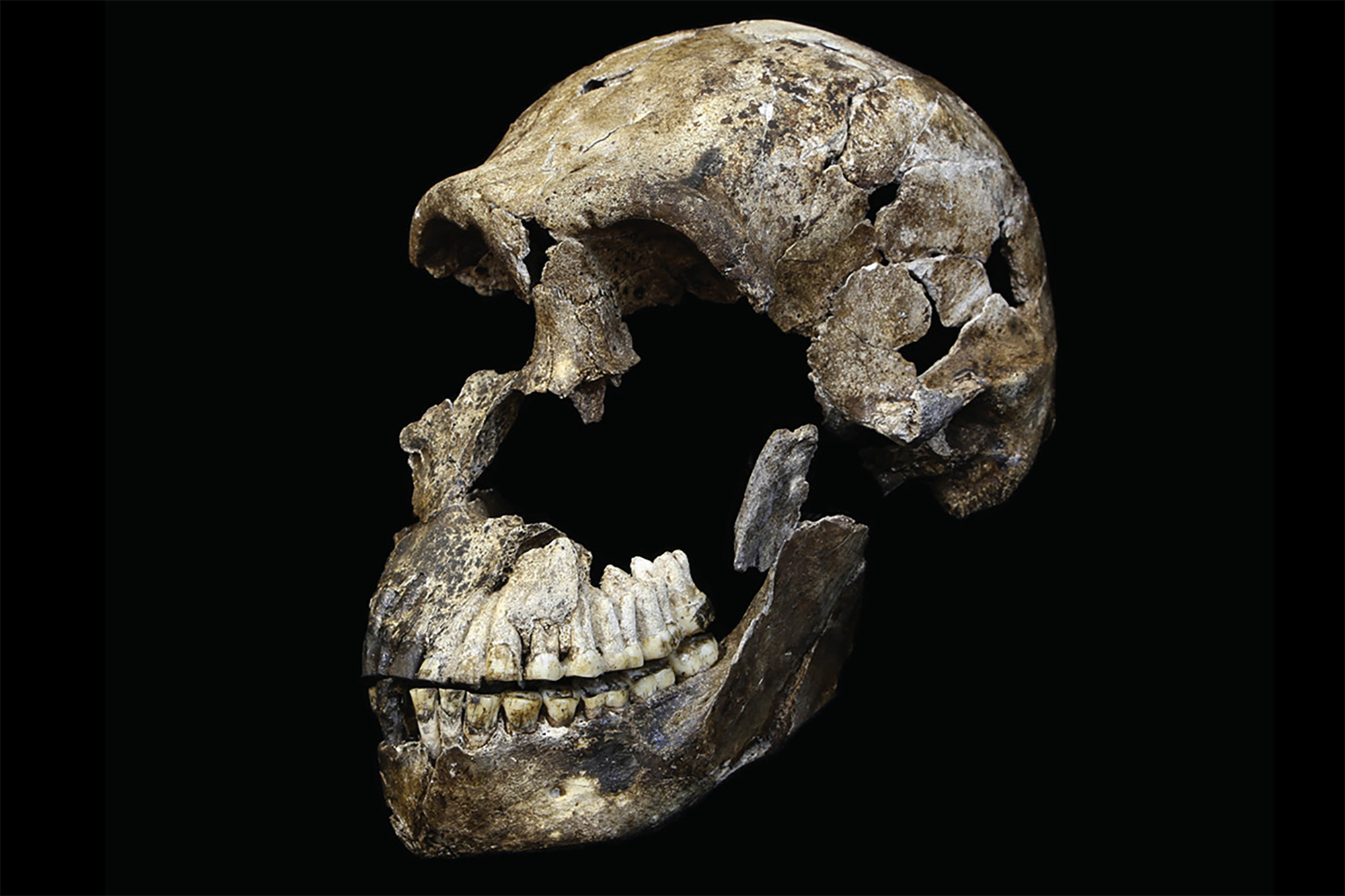
Two male H. naledi skulls from the Dinaledi chamber had cranial volumes of about 560 cm3 (34 cu in), and two female skulls 465 cm3 (28.4 cu in). A male H. naledi skull from the Lesedi chamber had a cranial volume of 610 cm3 (37 cu in). The Dinaledi specimens are more similar to the cranial capacity of australopithecines; for comparison, H. erectus averaged about 900 cm3 (55 cu in), and modern humans 1,270 and 1,130 cm3 (78 and 69 cu in) for males and females respectively. The Lesedi specimen is more within the range of H. habilis and H. e. georgicus. Nonetheless, the encephalization quotient of H. naledi was estimated at 3.75, which is the same as the pygmy H. floresiensis, but notably smaller than all other Homo. Contemporary Homo were all above 6, H. e. georgicus at 3.55, and A. africanus at 3.81. It is unclear if H. naledi inherited small brain size from the last common Homo ancestor, or if it was evolved secondarily more recently.
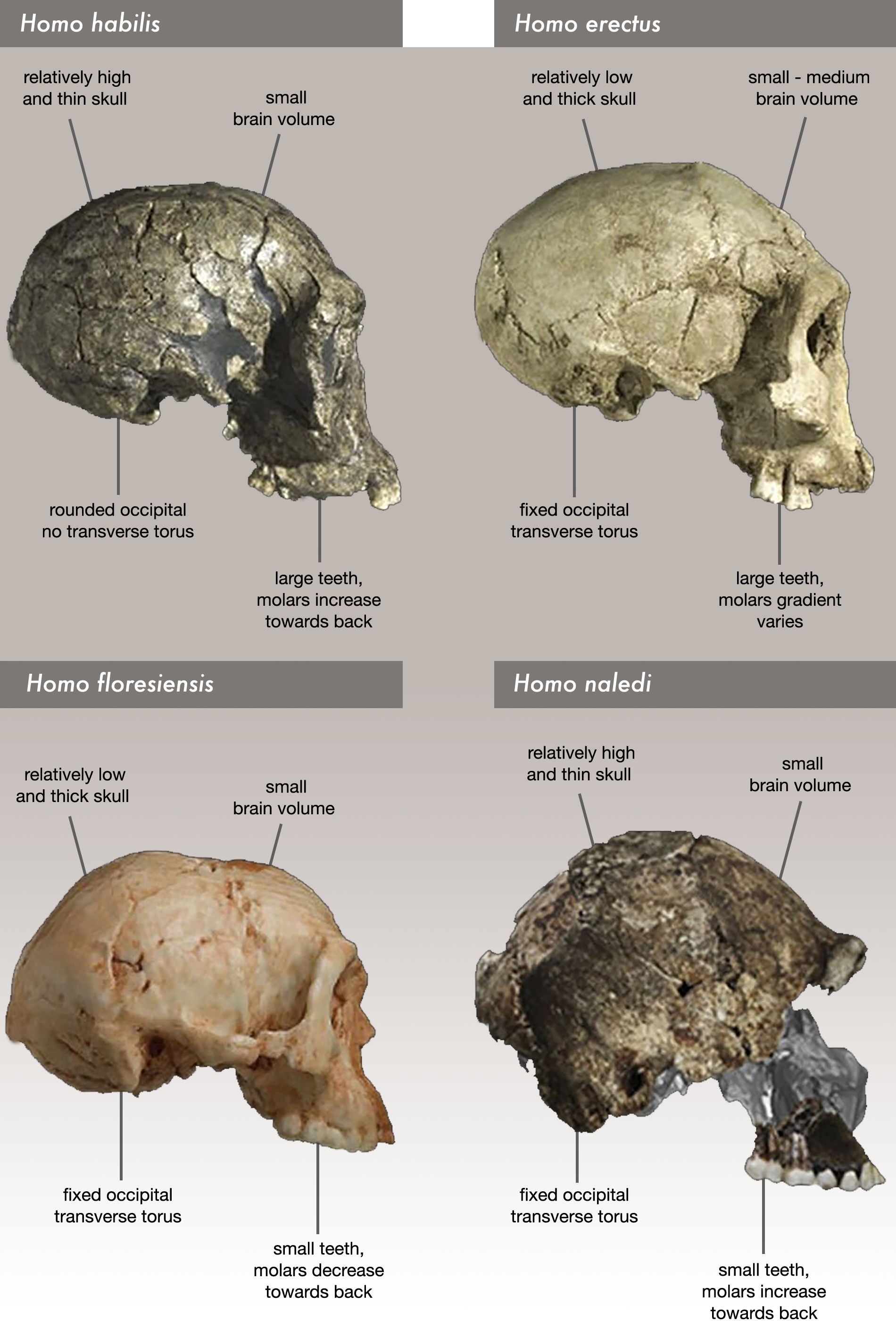
Like modern humans, but unlike fossil hominins (including South African australopithecines, H. erectus, and Neanderthals), the permanent 2nd molar erupted comparatively late in life, emerging alongside the premolars instead of before, which indicates a slower maturation unusually comparable to modern humans. The tooth formation rate of the front teeth is also most similar to modern humans. The overall size and shape of the molars most closely resemble those of three unidentified Homo specimens from the local Swartkrans and East African Koobi Fora Caves, and are similar in size (but not shape) to Pleistocene H. sapiens. The necks of the molars are proportionally similar to those of A. afarensis and Paranthropus. Unlike modern humans and contemporary Homo, H. naledi lacks several accessory dental features, and has a high frequency of individuals who present main cusps, namely the metacone (midline on the tongue-side) and hypocone (to the right on the lip-side) on the 2nd and 3rd molars, and a Y-shaped hypoconulid (a ridge on the lip-side towards the cheek) on all 3 molars. Nonetheless, H. naledi also has many dental similarities with contemporary Homo.
The anvil (a middle ear bone) more resembles those of chimps, gorillas, and Paranthropus than Homo. Like H. habilis and H. erectus, H. naledi has a well-developed brow-ridge with a fissure stretching across just above the ridge, and like H. erectus a pronounced occipital bun. H. naledi has some facial similarities with H. rudolfensis.
Build
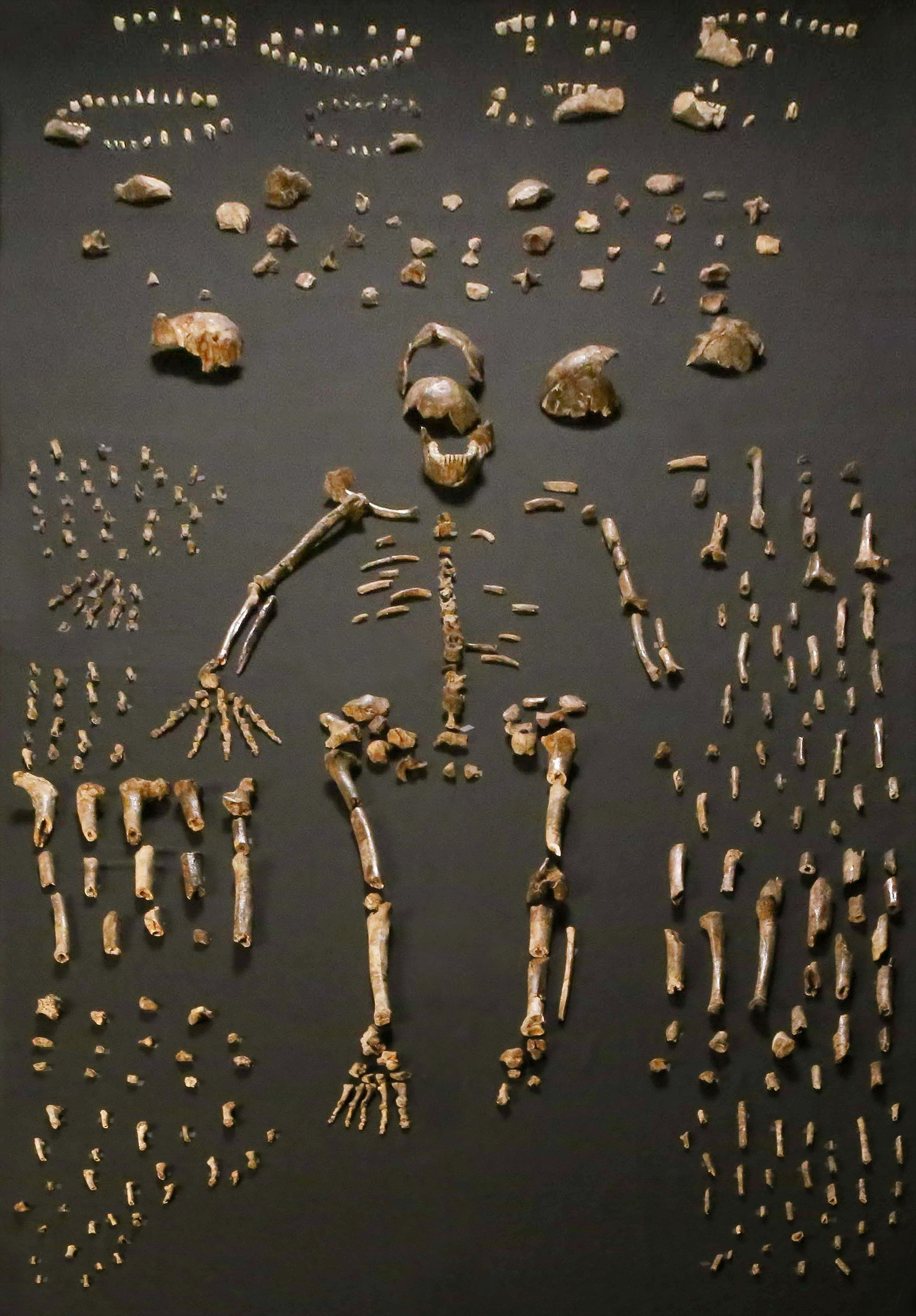
Concerning the spine, only the 10th and 11th thoracic vertebrae (in the chest region) are preserved from presumably a single individual, which are proportionally similar to those of contemporary Homo, though are the smallest recorded of any hominin. The two transverse processes of the vertebra, which jut out diagonally, are most similar to those of Neanderthals. The neural canals within are proportionally large, similar to modern humans, Neanderthals, and H. e. georgicus. The 11th rib is straight like that of A. afarensis, and the 12th rib is robust in cross-section like that of Neanderthals. Like Neanderthals, the 12th rib appears to have supported strong intercostal muscles above, and a strong quadratus lumborum muscle below. However, unlike Neanderthals, there was weak attachment to the diaphragm. Overall, this H. naledi specimen appears to have been small-bodied compared to other Homo, though it is unclear if this single specimen is representative of the species.
The shoulders are more similar to those of australopithecines, with the shoulder blade situated higher on the back and farther from the midline, short clavicles, and little or no humeral torsion. Elevated shoulder and clavicle bones indicate a narrow chest. The pelvis and legs have features reminiscent of Australopithecus, including anterposteriorly compressed (from front to back) femoral necks, mediolaterally compressed (from left to right) tibiae, and a somewhat circular fibular neck; which indicate a wide abdomen. This combination would preclude efficient endurance running in H. naledi, unlike H. erectus and descendants. Instead, H. naledi appears to have been more arboreal.
Limbs
The metacarpal bone of the thumb was well-developed, which is used in holding and manipulating large objects, and had strong crests to support its opponens pollicis muscle used in precision-pinch gripping, and its thenar muscles. This is more similar to other Homo than Australopithecus. H. naledi appears to have had strong flexor pollicis longus muscles like modern humans, with humanlike palm and finger pads, which are important in forceful gripping between the thumb and fingers. However, unlike Homo, the H. naledi thumb metacarpal joint is comparably small relative to the thumb's length, and the thumb phalangeal joint is flattened. The distal thumb phalanx bone is robust, and proportionally more similar to those of H. habilis and P. robustus.
The metacarpals of the other fingers share adaptations with modern humans and Neanderthals to cup and manipulate objects, and the wrist joint is overall comparable to that of modern humans and Neanderthals. Conversely, the proximal phalanges are curved and are almost identical to those of A. afarensis and H. habilis, which is interpreted as an adaptation for climbing and suspensory behaviour. Such curvature is more pronounced in adults than juveniles, suggesting that adults climbed just as much or more so than juveniles, and this behaviour was commonly done. The fingers are also proportionally longer than those of any other fossil hominin (other than the arboreal Ardipithecus ramidus and a modern human specimen from Qafzeh cave, Israel) which is also consistent with climbing behaviour.
H. naledi was a biped and stood upright. Like other Homo, they had strong insertion for the gluteus muscles, well-defined linea aspera (a ridge running down the back of the femur), thick patellae, long tibiae, and gracile fibulae. These indicate that they were capable of long-distance travel. The H. naledi foot was similar to that of modern humans and other Homo, with adaptations for bipedalism and a humanlike gait. However, the heel bone has a low orientation, comparable to those of non-human great apes, and the ankle bone has a low declination, which possibly indicate the foot would have been subtly stiffer during the stance phase of walking before the foot pushed off the ground.
Pathology
The adult right mandible U.W. 101-1142 has a bony lesion, suggestive of a benign tumour. The individual would have experienced some swelling and localised discomfort, but the tumour's position near the medial pterygoid muscle may have impeded function of the muscle, and changed elevation of the right side of the jaw, and caused discomfort on the right temporomandibular joint (connecting the jaw with the skull).
Dental defects in H. naledi specimens during 1.6–2.8 and 4.3–7.6 months of development were most likely caused by seasonal stressors. This may have been due to extreme summer and winter temperatures causing food scarcity. Also, minimum winter temperatures of the area average about 3 °C (37 °F), and can drop below freezing; staying warm for an infant of the small-bodied H. naledi would have been difficult, and winters likely increased susceptibility to respiratory diseases. Environmental stressors is consistent with present-day flu seasons in South Africa peaking during winter, and paediatric diarrhoea hospitalisation being most frequent at the height of the rainy season in summer.
Local hominins were likely preyed upon by large carnivores, such as lions, leopards, and hyaenas. However, there seems to be a distinct paucity of large carnivore remains from the northern end of the Cradle of Humankind, where Rising Star Cave is located, possibly because carnivores preferred the Blaaubank River to the south which may have offered better hunting grounds with a greater abundance of large prey items. Alternatively, because many more sites are known in the south than the north, carnivore spatial patterns may not be well-represented by the fossil record.
Food
Dental chipping and wearing indicates the habitual consumption of small hard objects, such as dirt and dust, and cup-shaped wearing on the back teeth may have stemmed from gritty particles. These could have originated from unwashed roots and tubers. Alternatively, aridity could have stirred up particulates onto food items, coating food in dust. It is possible that they commonly ate larger hard items, such as seeds and nuts, but these were processed into smaller pieces before consumption. Whatever the case, H. naledi occupied a seemingly unique niche from previous South African hominins, including Australopithecus and Paranthropus. However, the teeth of all 3 species indicate that they needed to exert high shearing force to chew through perhaps plant or muscle fibres. The teeth of other Homo cannot produce such high forces perhaps due to the use of some food processing techniques, such as cooking.
Technology
Though H. naledi remains are not associated with any stone tools, it is likely they produced Early Stone Age (Acheulean and possibly the earlier Oldawan) or Middle Stone Age industries because they have the same adaptations to the hand as other human species which are implicated in tool production. H. naledi is the only identified human species to have existed during the early Middle Stone Age of the Highveld region, South Africa, possibly indicating that this species manufactured and maintained this tradition at least during this time period. Such industries and stone cutting techniques likely evolved independently several times among different Homo species and populations, or were transported over long distances by the inventors or apprentices and taught.