Homo sapiens
Hominin Context Quick Guide



Homo sapiens
Hominin traits Archaeological industry/Technocomplex including art
Period in human prehistory: M = Mesolithic; N = Neolithic; B = Bronze Age; I = Iron Age; Geological epoch
* Note: Table based past and current research and scientific consensus
Hominin traits Archaeological industry/Technocomplex including art
Period in human prehistory: M = Mesolithic; N = Neolithic; B = Bronze Age; I = Iron Age; Geological epoch
* Note: Table based past and current research and scientific consensus
Homo sapiens
Hominin traits
Archaeological industry/Technocomplex including art
Aur = Aurignacian; Mag = Magdalenian;
Grav = Gravettian; Sol = Solutrean
Period in human prehistory:
M = Mesolithic; N = Neolithic;
B = Bronze Age; I = Iron Age;
Geological epoch
* Note: Table based past and current research
and scientific consensus
Hominin traits
Archaeological industry/Technocomplex including art
Aur = Aurignacian; Mag = Magdalenian;
Grav = Gravettian; Sol = Solutrean
Period in human prehistory:
M = Mesolithic; N = Neolithic;
B = Bronze Age; I = Iron Age;
Geological epoch
* Note: Table based past and current research
and scientific consensus
Homo sapiens are primates of the Family Hominidae and are the only living species of their genus. This lineage originated 200 thousand years ago in Eastern Africa and have since spread to all corners of the world.
The Origins of Modern Humans
by Cassandra Turcotte
Center for the Advanced Study of Hominid Paleobiology (CASHP)
Center for the Advanced Study of Hominid Paleobiology (CASHP)

Several lines of evidence support a single African origin for Homo sapiens. For example, analysis of mitochondrial DNA has demonstrated that the greatest level of genetic diversity in modern humans occurs within the African continent, represented by the mtDNA macro-haplogroup L (Ingman et al. 2000). Mitochondrial DNA, of course, is passed from mother to daughter without recombination and thereby preserves a record of female descent to the matrilineal most recent common ancestor - the woman from whom all humans alive today descend (see Pakendorf and Stoneking 2005; van Oven and Kayser 2008).
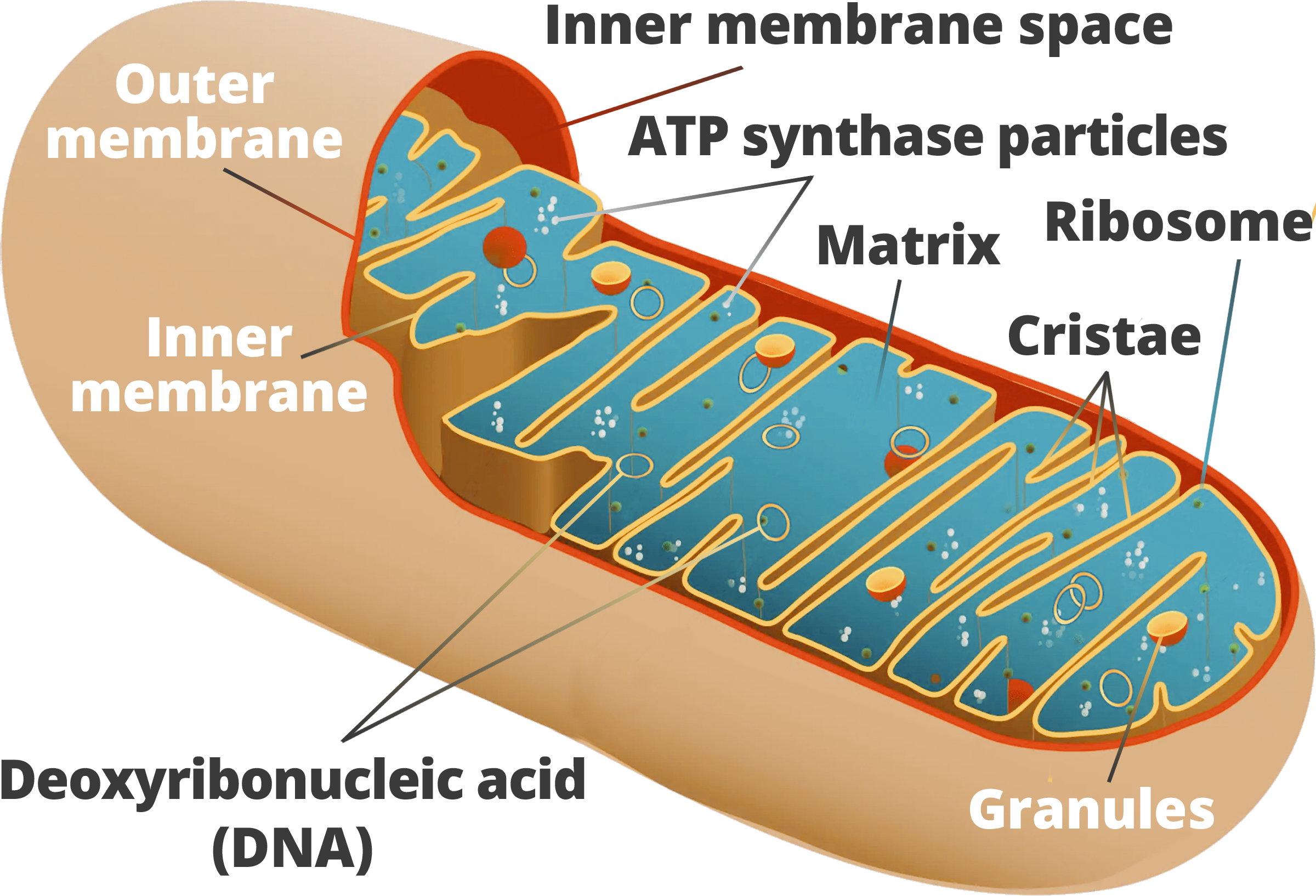
Mitochondrial DNA has demonstrated that the greatest level of genetic diversity in modern humans occurs within the African continent
This woman, nicknamed Mitochondrial Eve, lived some time between 99 and 200 kyr in East Africa (Cann et al. 1987; Ayala 1995; Jorde et al. 1998). Her descendants gave rise to all present-day variation. Today, there are seven primary subclades of the L haplogroup (L0-L6), all of which present primarily in Africa. Diversity in all non-Africans, including Europeans, Asians and Native Americans, is a subset of the L3 haplogroup (Jorde et al. 1998; van Oven and Kayser 2008).
The earliest human fossils also come from Africa. More specifically, these fossils are found in Ethiopia. The fossil skulls at Omo Kibish attributed to modern Homo sapiens (Day 1969) date all the way back to ~195 kyr (McDougall et al. 2005). Additional finds from Herto Bouri, Ethiopia, dated to ~160 kyr, were characterized as morphologically primitive anatomically modern humans (White et al. 2003; Clark et al. 2003), which the authors refer to as Homo sapiens idaltu to differentiate them from later Homo sapiens sensu stricto. Both are part of the same species, though earlier Homo sapien idaltu is thought to preserve more archaic morphology such as increased robusticity, prognathism and lack of chin (White et al. 2003; McBrearty and Brooks 2000). H. sapiens are defined in contrast to earlier specimens by a more globular cranium, prevailing orthognathism, and skeletal gracility on top of behavioral modernity (McBrearty and Brooks 2000).
Homo sapiens idaltu

The fossilized remains of Homo sapiens idaltu were discovered in 1997 by Tim White at Herto Bouri near the Middle Awash site of Ethiopia’s Afar Triangle. Dating took place by the radioisotope method which analysed the volcanic layers containing the 3 cranial fossils [White 2003].
The morphology of the skulls display archaic features not found in the later Homo sapiens, but are still seen as the direct ancestors of modern Homo sapiens sapiens.
The remains discovered at Herto Bouri have been named 'Herto Man'. Experts claim the finds are complete enough to be identified as early modern humans, since they show the characteristic globular shape of the braincase and the facial features of our species. However, both the adult skulls are huge and robust, and also show resemblances to more primitive African fossils.
Expansion
| HOMO SAPIENS / HOMO SAPIENS SAPIENS | |
 | |
| Genus: | Homo |
| Species: | Homo sapiens / sapiens sapiens |
| Other Names: | • Modern Humans • Cro-Magnon • Omo |
| Time Period: |
• Homo sapiens • 200,000 - 50,000 years ago • Homo sapiens sapiens • 50,000 years ago to Present |
| Characteristics: | Modern, Migration, Art, More Gracile, Technological |
| Fossil Evidence: | Fossil Specimens, Europe, Worldwide |

Anatomical comparison of the skulls of Homo sapiens sapiens (modern humans) and Homo neanderthalensis (right)
Skeletal evidence places modern humans in Australia possibly ca. 60 kyr in the Lake Mungo region (Stringer 1999; Thorne et al. 1999; Balme 2013; Long et al. 2014), and in China’s Tianyuan Cave by 40 kyr (Tong et al. 2004; Shang and Trinkaus 2010). Definitive evidence of modern humans in Central Asia and Europe, however, is complicated by a fragmentary fossil record and the presence of similar-looking Neanderthal specimens. Recent work has identified ~45 kyr molars from Grotta del Cavallo in Italy as the first such remains of Homo sapiens in the region (Benazzi et al. 2011), followed by revised dates for the Kent’s Cavern, UK modern humans, placed to ~44 kyr (Higham et al. 2011). Settlement of the Americas occurred relatively late in the scheme of human evolution, delayed by the physical barrier of the Pacific Ocean until glacial periods lowered the sea level enough to expose a land bridge connecting modern day Siberia to Alaska (See Achilli et al. 2013; Meiri et al. 2014; Raff and Bolnick 2014).
An Out of Africa perspective on modern human origins and expansion contrasts what is known as the Multiregional model, where humans are believed to have evolved from specimens of ≈in situ around the world. In this way, regional differences accrued but species cohesion was maintained through genetic drift (Stringer and Andrews 1988; Wolpoff et al. 1994). This model expects transitional fossils outside of Africa and regional continuity in morphology. While there is some evidence of genetic admixture (Currat and Excoffier 2011; Prüfer et al. 2014), the long genetic and fossil history of H. sapiens in Africa paints a strong Multiregional view of modern human origins as problematic and the Out of Africa model a better fit of current evidence.
Behavioral Modernity

Cave paintings of Lascaux in France
Traditionally, modern human behavior was thought to arise suddenly and at once across Europe 40-50 kyr, implying quite a long lag between anatomical and behavioral modernity. On top of unfairly portraying preceding human populations as primitive, however, this view is inaccurate. Indeed, the components of behavioral modernity appear within the African Middle Stone Age (McBrearty and Brooks 2000). These activities represent manifestations of advanced cognitive behaviors: abstract thinking, planning, innovation, an appreciation of symbolism.

Venus of Dolní Věstonice

Venus of Dolní Věstonice
Of course, humans still exist today, pushing the envelope of cultural and technological evolution to ever more spectacular limits. The physical and behavioral modernity of this species have their roots in Africa and subsequently spread around the globe, replacing earlier hominins either through competition or hybridization. The implications of a single successful exit are highly significant, and lie at the core of the Journey of Mankind Genetic Map.
The question of course is now ‘will the Homo genus keep evolving?’ Moreover, what other discoveries will help explain the ‘untidy tree’ of evolution?
References
Achilli A, Perego UA, Lancioni H, Olivieri A, Gandini F, Kashani BH, Battaglia V, Grugni V, Angerhofer N, Rogers MP, Herrera RJ, Woodward SR, Labuda D, Smith DG, Cybulski JS, Semino O, Malhi RS and A Torroni (2013) Reconciling migration models to the Americas with variation of North American native mitogenomes. PNAS 110.35: 1-6.
Ambrose SH (1998) Chronology of the Later Stone Age and food production in East Africa. J Archaeol Sci 25: 377-392.
Ayala FJ (1995) The myth of Eve: molecular biology and human origins. Science 270:263-291.
Balme J (2013) Of boats and string: the maritime colonisation of Australia. Quat Int 285: 68-75.
Behar DM, Villems R, Soodyal H, Blue-Smith J, Pereira L, Metspalu E, Scozzari R, Makkan H, Tzur S, Comas D, Bertranpetit J, Quintana-Murci L, Tyler-Smith C, Wells RS and S Rosset (2008) The dawn of human matrilineal diversity. Cell Press 82.5: 1130-1140.
Benazzi S, Douka K, Fornai C, Bauer CC, Kullmer O, Svoboda J, Pap I, Mallegni F, Bayle P, Coquerelle M, Condemi S, Ronchitelli A, Havarti K and GW Weber (2011) Early dispersal of modern humans in Europe and implications for Neanderthal behavior. Nature 479: 525-528.
Blackwell L, d’Errico F and L Wadley (2008) Middle Stone Age bone tools from the Howiesoons Poort layers, Sibudu Cave, South Africa. J Archaeol Sci 35.6: 1566-1580.
Brooks AS (1995) L’Aurignacien de l’abri Pataud niveaux 6 a 14. Ed. H. M. Bricker. In Le Paléolithique Supérieur de l’Abri Pataud (Dordogne): Les Fouilles de H. L. Movius, Jr. Paris: Documents d’Archéologie Française, Maison des Sciences de l’Homme.
Cann RL (1988) DNA and human origins. A. Rev. Anthropol 17: 127-143.
Clark JD, Beyene Y, WoldeGabriel G, Hart WK, Renne PR, Gilbert H, Defleur A, Suwa G, Katoh S, Ludwig KR, Boisserie J-R, Asfaw B and TD White (2003) Stratigraphic, chronological and behavioral contexts of Currat M and L Excoffier (2011) Strong reproductive isolation between humans and Neanderthals inferred from observed patterns of introgression. Proc Natl Acad Sci 108: 15129-15132.
Day MH (1969) Omo human skeletal remains. Nature 222: 1135-1138.
Debénath A (1994) L’Atérien du nord de l’Afrique du Sahara. Sahara 6: 21-30.
Goodwin AJH (1929) An introduction to the Middle Stone Age in South Africa. S Afr J Sci 25: 410-418.
Henshilwood C and JC Sealey (1997) Bone artefacts from the Middle Stone Age at Blombos Cave, Southern Cape, South Africa. Curr Anthropol 38: 890-895.
Higham T, Compton T, Stringer C, Jacobi R, Shapiro B, Trinkaus E and B Chandler (2011) The earliest evidence for anatomically modern humans in northwestern Europe. Nature 479.7374: 521-524.
Ingman M, Kaessmann H, Pääbo S and U Gyllensten (2000) Mitochondrial genome variation and the origin of modern humans. Nature 408: 708-713.
Jorde LB, Bamshad M and AR Rogers (1998) Using mitochondrial and nuclear DNA markers to reconstruct human evolution. BioEssays 20.2: 126-136.
Klein RG (1994) Southern Africa before the Iron Age in Integrative Pathways to the Past: Paleoanthropological investigations in honor of F. Clark Howell. Eds. RS Corrucini and RL Ciochon. Prentice Hall, New Jersey.
Kokis JE, Hare PE and AS Brooks (1998) Stratigraphic analysis of White Paintings Shelter, Botswana, using isoleucine epimerization in ratite eggshell. Amino Acids 15: 280-281.
Leakey MD (1971) Olduvai Gorge, Vol. 3: Excavations in Beds I and II, 1960-1963. Cambridge: Cambridge University Press.
Long K, Stern N, Williams IS, Kinsley L, Wood R, Sporcic K, Smith T, Fallon S, Kokkonen H, Moffat I and R Grün (2014) Fish otolith geochemistry, environmental conditions and human occupation at Lake Mungo, Australia. Quat Sci Rev 88: 82-95.
McBrearty S and AS Brooks (2000) The revolution that wasn’t: A new interpretation of modern human behavior. J Hum Evol 39: 453-563.
McDougall I, Brown FH and JG Fleagle (2005) Stratigraphic placement and age of modern humans from Kibish, Ethiopia. Nature 433: 733-736.
Meiri M, Lister AM, Collins MJ, Tuross N, Goebel T, Blockley S, Zazula GD, van Doorn N, Guthrie RD, Boeskorov GG, Baryshnikov GF, Sher A and I Barnes (2014) Faunal record identifies Bering isthmus conditions as constraint to end-Pleistocene migration to the New World. Proc R Soc B 281: 1-9.
Milo R (1998) Evidence for hominid predation at Klasies River Mouth, South Africa, and its implications for the behavior of early modern humans. J Archaeol Sci 25: 99-133.
Pakendorf B and M Stoneking (2005) Mitochondrial DNA and human evolution. Annu Rev Genomics Hum Genet 6: 165-183.
Prüfer K, Racimo F, Patterson N, Jay F, Sankararaman S, Sawyer S, Heinze A, Renaud G, Sudmant PH, de Filippo C, Li H, Mallick S, Dannemann M, Fu Q, Kircher M, Kuhlwilm M, Lachmann M, Meyer M, Ongyerth M, Siebauer M, Theunert C, Tandon A, Moorjani P, Pickrell J, Mullikin JC, Vohr SH, Green RE, Hellmann I, Johnson PLF, Blanche H, Cann H, Kitzman JO, Shendure J, Eichler EE, Lein ES, Bakken TE, Golovanova L, Doronichev VB, Shunkov MV, Derevianko AP, Viola B, Slatkin M, Reich D, Kelso J and S Pääbo (2014) The complete genome sequence of a Neanderthal from the Altai Mountains. Nature 505: 43-49.
Raff JA and DA Bolnick (2014) Palaeogenomics: Genetic roots of the first Americans. Nature 506: 162-163.
Robbins et al. (1994) Barbed bone points, paleoenvironment and the antiquity of fish exploitation in the Kalahari Desert, Botswana. J Field Archaeol 21: 257-264.
Robbins LH, Murphy ML, Brooks GA, Ivester A, Campbell AC, Klein RG, Milo R, Brooks AS, Kokis JE and W Downey (1999) Archaeology, paleoenvironment and chronology of the White Paintings Rock Shelter, Tsodilo Hills, northwest Kalahari Desert, Botswana. J. Archaeol Sci.
Sankararaman S, Patterson N, Li H, Pääbo S and D Reich (2012) The date of interbreeding between Neanderthals and modern humans. PLoS Genetics 8.10.
Sankararaman S, Mallick S, Dannemann M, Prüfer K, Kelso J, Pääbo S, Patterson N and D Reich (2014) The genomic landscape of Neanderthal ancestry in present-day humans. Nature 507: 354-357.
Shang H and E Trinkaus (2010) The early modern human from Tianyuan Cave. Texas A&M University Press: College Station, TX.
Singer R and J Wymer (1982) The Middle Stone Age at Klasies River Mouth in South Africa. Chicago: University of Chicago Press.
Soares P, Alshamali F, Pereira JB, Fernandes V, Silva NM, Afonso C, Costa MD, Musilova, Macaulay V, Richards MB, Cerny V and L Pereira (2012) The expansion of mtDNA haplogroup L3 within and out of Africa. Mol Biol Evol 29.3: 915-927.
Stringer CB and P Andrews (1988) Geneetic and fossil evidence for the origin of modern humans. Science 239: 1263-1268.
Stringer CB (1999) Has Australia backdated the Human Revolution? Antiquity 73: 876-879.
Stringer C (2001) Modern human origins: progress and prospects. Phil Trans R Soc Lond B 357: 563-597.
Thorne A , Grun R, Mortimer B, Spooner N, Simpson J, McCulloch M, Taylor L and D Curnoe (1999) Australia’s oldest human remains: age of the Lake Mungo 3 skeleton. J Hum Evol 36: 591-612.
Tong HW, Shang H, Zhang SQ and FY Chen (2004) A preliminary report on the newly found Tianyuan Cave, a Late Pleistocene human fossil site near Zhoukoudian. Chin Sci Bull 49.8: 853-857.
van Oven M and M Kayser (2008) Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation. Human Mutation 1039: 386-394.
White TD, Asfaw B, DeGusta D, Gilbert H, Richards GD, Suwa G and FC Howell (2003) Pleistocene Homo sapiens from Middle Awash Ethiopia. Nature 423: 742-747.
Wolpoff M, Thorne AG, Smith FH, Frayer DW and GG Pope (1994) Multiregional evolution: A world-wide source for modern human populations in Origins of anatomically modern humans. Springer, USA.
Ambrose SH (1998) Chronology of the Later Stone Age and food production in East Africa. J Archaeol Sci 25: 377-392.
Ayala FJ (1995) The myth of Eve: molecular biology and human origins. Science 270:263-291.
Balme J (2013) Of boats and string: the maritime colonisation of Australia. Quat Int 285: 68-75.
Behar DM, Villems R, Soodyal H, Blue-Smith J, Pereira L, Metspalu E, Scozzari R, Makkan H, Tzur S, Comas D, Bertranpetit J, Quintana-Murci L, Tyler-Smith C, Wells RS and S Rosset (2008) The dawn of human matrilineal diversity. Cell Press 82.5: 1130-1140.
Benazzi S, Douka K, Fornai C, Bauer CC, Kullmer O, Svoboda J, Pap I, Mallegni F, Bayle P, Coquerelle M, Condemi S, Ronchitelli A, Havarti K and GW Weber (2011) Early dispersal of modern humans in Europe and implications for Neanderthal behavior. Nature 479: 525-528.
Blackwell L, d’Errico F and L Wadley (2008) Middle Stone Age bone tools from the Howiesoons Poort layers, Sibudu Cave, South Africa. J Archaeol Sci 35.6: 1566-1580.
Brooks AS (1995) L’Aurignacien de l’abri Pataud niveaux 6 a 14. Ed. H. M. Bricker. In Le Paléolithique Supérieur de l’Abri Pataud (Dordogne): Les Fouilles de H. L. Movius, Jr. Paris: Documents d’Archéologie Française, Maison des Sciences de l’Homme.
Cann RL (1988) DNA and human origins. A. Rev. Anthropol 17: 127-143.
Clark JD, Beyene Y, WoldeGabriel G, Hart WK, Renne PR, Gilbert H, Defleur A, Suwa G, Katoh S, Ludwig KR, Boisserie J-R, Asfaw B and TD White (2003) Stratigraphic, chronological and behavioral contexts of Currat M and L Excoffier (2011) Strong reproductive isolation between humans and Neanderthals inferred from observed patterns of introgression. Proc Natl Acad Sci 108: 15129-15132.
Day MH (1969) Omo human skeletal remains. Nature 222: 1135-1138.
Debénath A (1994) L’Atérien du nord de l’Afrique du Sahara. Sahara 6: 21-30.
Goodwin AJH (1929) An introduction to the Middle Stone Age in South Africa. S Afr J Sci 25: 410-418.
Henshilwood C and JC Sealey (1997) Bone artefacts from the Middle Stone Age at Blombos Cave, Southern Cape, South Africa. Curr Anthropol 38: 890-895.
Higham T, Compton T, Stringer C, Jacobi R, Shapiro B, Trinkaus E and B Chandler (2011) The earliest evidence for anatomically modern humans in northwestern Europe. Nature 479.7374: 521-524.
Ingman M, Kaessmann H, Pääbo S and U Gyllensten (2000) Mitochondrial genome variation and the origin of modern humans. Nature 408: 708-713.
Jorde LB, Bamshad M and AR Rogers (1998) Using mitochondrial and nuclear DNA markers to reconstruct human evolution. BioEssays 20.2: 126-136.
Klein RG (1994) Southern Africa before the Iron Age in Integrative Pathways to the Past: Paleoanthropological investigations in honor of F. Clark Howell. Eds. RS Corrucini and RL Ciochon. Prentice Hall, New Jersey.
Kokis JE, Hare PE and AS Brooks (1998) Stratigraphic analysis of White Paintings Shelter, Botswana, using isoleucine epimerization in ratite eggshell. Amino Acids 15: 280-281.
Leakey MD (1971) Olduvai Gorge, Vol. 3: Excavations in Beds I and II, 1960-1963. Cambridge: Cambridge University Press.
Long K, Stern N, Williams IS, Kinsley L, Wood R, Sporcic K, Smith T, Fallon S, Kokkonen H, Moffat I and R Grün (2014) Fish otolith geochemistry, environmental conditions and human occupation at Lake Mungo, Australia. Quat Sci Rev 88: 82-95.
McBrearty S and AS Brooks (2000) The revolution that wasn’t: A new interpretation of modern human behavior. J Hum Evol 39: 453-563.
McDougall I, Brown FH and JG Fleagle (2005) Stratigraphic placement and age of modern humans from Kibish, Ethiopia. Nature 433: 733-736.
Meiri M, Lister AM, Collins MJ, Tuross N, Goebel T, Blockley S, Zazula GD, van Doorn N, Guthrie RD, Boeskorov GG, Baryshnikov GF, Sher A and I Barnes (2014) Faunal record identifies Bering isthmus conditions as constraint to end-Pleistocene migration to the New World. Proc R Soc B 281: 1-9.
Milo R (1998) Evidence for hominid predation at Klasies River Mouth, South Africa, and its implications for the behavior of early modern humans. J Archaeol Sci 25: 99-133.
Pakendorf B and M Stoneking (2005) Mitochondrial DNA and human evolution. Annu Rev Genomics Hum Genet 6: 165-183.
Prüfer K, Racimo F, Patterson N, Jay F, Sankararaman S, Sawyer S, Heinze A, Renaud G, Sudmant PH, de Filippo C, Li H, Mallick S, Dannemann M, Fu Q, Kircher M, Kuhlwilm M, Lachmann M, Meyer M, Ongyerth M, Siebauer M, Theunert C, Tandon A, Moorjani P, Pickrell J, Mullikin JC, Vohr SH, Green RE, Hellmann I, Johnson PLF, Blanche H, Cann H, Kitzman JO, Shendure J, Eichler EE, Lein ES, Bakken TE, Golovanova L, Doronichev VB, Shunkov MV, Derevianko AP, Viola B, Slatkin M, Reich D, Kelso J and S Pääbo (2014) The complete genome sequence of a Neanderthal from the Altai Mountains. Nature 505: 43-49.
Raff JA and DA Bolnick (2014) Palaeogenomics: Genetic roots of the first Americans. Nature 506: 162-163.
Robbins et al. (1994) Barbed bone points, paleoenvironment and the antiquity of fish exploitation in the Kalahari Desert, Botswana. J Field Archaeol 21: 257-264.
Robbins LH, Murphy ML, Brooks GA, Ivester A, Campbell AC, Klein RG, Milo R, Brooks AS, Kokis JE and W Downey (1999) Archaeology, paleoenvironment and chronology of the White Paintings Rock Shelter, Tsodilo Hills, northwest Kalahari Desert, Botswana. J. Archaeol Sci.
Sankararaman S, Patterson N, Li H, Pääbo S and D Reich (2012) The date of interbreeding between Neanderthals and modern humans. PLoS Genetics 8.10.
Sankararaman S, Mallick S, Dannemann M, Prüfer K, Kelso J, Pääbo S, Patterson N and D Reich (2014) The genomic landscape of Neanderthal ancestry in present-day humans. Nature 507: 354-357.
Shang H and E Trinkaus (2010) The early modern human from Tianyuan Cave. Texas A&M University Press: College Station, TX.
Singer R and J Wymer (1982) The Middle Stone Age at Klasies River Mouth in South Africa. Chicago: University of Chicago Press.
Soares P, Alshamali F, Pereira JB, Fernandes V, Silva NM, Afonso C, Costa MD, Musilova, Macaulay V, Richards MB, Cerny V and L Pereira (2012) The expansion of mtDNA haplogroup L3 within and out of Africa. Mol Biol Evol 29.3: 915-927.
Stringer CB and P Andrews (1988) Geneetic and fossil evidence for the origin of modern humans. Science 239: 1263-1268.
Stringer CB (1999) Has Australia backdated the Human Revolution? Antiquity 73: 876-879.
Stringer C (2001) Modern human origins: progress and prospects. Phil Trans R Soc Lond B 357: 563-597.
Thorne A , Grun R, Mortimer B, Spooner N, Simpson J, McCulloch M, Taylor L and D Curnoe (1999) Australia’s oldest human remains: age of the Lake Mungo 3 skeleton. J Hum Evol 36: 591-612.
Tong HW, Shang H, Zhang SQ and FY Chen (2004) A preliminary report on the newly found Tianyuan Cave, a Late Pleistocene human fossil site near Zhoukoudian. Chin Sci Bull 49.8: 853-857.
van Oven M and M Kayser (2008) Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation. Human Mutation 1039: 386-394.
White TD, Asfaw B, DeGusta D, Gilbert H, Richards GD, Suwa G and FC Howell (2003) Pleistocene Homo sapiens from Middle Awash Ethiopia. Nature 423: 742-747.
Wolpoff M, Thorne AG, Smith FH, Frayer DW and GG Pope (1994) Multiregional evolution: A world-wide source for modern human populations in Origins of anatomically modern humans. Springer, USA.
Links
Origins Archive Index •
Origins Overview •
Short Story of Human Evolution •
13 Big Questions •
Stone Tools •
Origins Art
Homo
Homo sapiens • Denisova hominin • Homo floresiensis • Homo naledi • Homo neanderthalensis • Homo rhodesiensis • Homo helmei
Homo heidelbergensis • Homo antecessor • Homo cepranensis • Homo erectus • Homo erectus georgicus • Homo ergaster
Homo rudolfensis • Homo habilis • Homo gautengensis
Kenyanthropus
Kenyanthropus platyops
Paranthropus
Paranthropus robustus • Paranthropus boisei • Paranthropus aethiopicus
Australopithecus
Australopithecus sediba • Australopithecus garhi • Australopithecus africanus • Australopithecus bahrelghazali
Australopithecus afarensis • Australopithecus anamensis
Ardipithecus
Ardipithecus ramidus • Ardipithecus kadabba
Orrorin
Orrorin tugenensis
Sahelanthropus
Sahelanthropus tchadensis
Homo
Homo sapiens • Denisova hominin • Homo floresiensis • Homo naledi • Homo neanderthalensis • Homo rhodesiensis • Homo helmei
Homo heidelbergensis • Homo antecessor • Homo cepranensis • Homo erectus • Homo erectus georgicus • Homo ergaster
Homo rudolfensis • Homo habilis • Homo gautengensis
Kenyanthropus
Kenyanthropus platyops
Paranthropus
Paranthropus robustus • Paranthropus boisei • Paranthropus aethiopicus
Australopithecus
Australopithecus sediba • Australopithecus garhi • Australopithecus africanus • Australopithecus bahrelghazali
Australopithecus afarensis • Australopithecus anamensis
Ardipithecus
Ardipithecus ramidus • Ardipithecus kadabba
Orrorin
Orrorin tugenensis
Sahelanthropus
Sahelanthropus tchadensis
Origins Archive Index •
Origins Overview •
Short Story of Human Evolution
13 Big Questions • Stone Tools • Origins Art Homo
Homo sapiens • Denisova hominin • Homo floresiensis • Homo naledi
Homo neanderthalensis • Homo rhodesiensis • Homo helmei • Homo heidelbergensis
Homo antecessor • Homo cepranensis • Homo erectus • Homo erectus georgicus
Homo ergaster • Homo rudolfensis • Homo habilis • Homo gautengensis
Kenyanthropus
Kenyanthropus platyops
Paranthropus
Paranthropus robustus • Paranthropus boisei • Paranthropus aethiopicus
Australopithecus
Australopithecus sediba • Australopithecus garhi • Australopithecus africanus
Australopithecus bahrelghazali • Australopithecus afarensis • Australopithecus anamensis
Ardipithecus
Ardipithecus ramidus • Ardipithecus kadabba
Orrorin
Orrorin tugenensis
Sahelanthropus
Sahelanthropus tchadensis
13 Big Questions • Stone Tools • Origins Art Homo
Homo sapiens • Denisova hominin • Homo floresiensis • Homo naledi
Homo neanderthalensis • Homo rhodesiensis • Homo helmei • Homo heidelbergensis
Homo antecessor • Homo cepranensis • Homo erectus • Homo erectus georgicus
Homo ergaster • Homo rudolfensis • Homo habilis • Homo gautengensis
Kenyanthropus
Kenyanthropus platyops
Paranthropus
Paranthropus robustus • Paranthropus boisei • Paranthropus aethiopicus
Australopithecus
Australopithecus sediba • Australopithecus garhi • Australopithecus africanus
Australopithecus bahrelghazali • Australopithecus afarensis • Australopithecus anamensis
Ardipithecus
Ardipithecus ramidus • Ardipithecus kadabba
Orrorin
Orrorin tugenensis
Sahelanthropus
Sahelanthropus tchadensis
Origins Archive Index
Origins Overview
Short Story of Human Evolution
13 Big Questions
Stone Tools
Origins Art
Homo
Homo sapiens
Denisova hominin
Homo floresiensis
Homo naledi
Homo neanderthalensis
Homo rhodesiensis
Homo helmei
Homo heidelbergensis
Homo antecessor
Homo cepranensis
Homo erectus
Homo erectus georgicus
Homo ergaster
Homo rudolfensis
Homo habilis
Homo gautengensis
Kenyanthropus
Kenyanthropus platyops
Paranthropus
Paranthropus robustus
Paranthropus boisei
Paranthropus aethiopicus
Australopithecus
Australopithecus sediba
Australopithecus garhi
Australopithecus africanus
Australopithecus bahrelghazali
Australopithecus afarensis
Australopithecus anamensis
Ardipithecus
Ardipithecus ramidus
Ardipithecus kadabba
Orrorin
Orrorin tugenensis
Sahelanthropus
Sahelanthropus tchadensis
Origins Overview
Short Story of Human Evolution
13 Big Questions
Stone Tools
Origins Art
Homo
Homo sapiens
Denisova hominin
Homo floresiensis
Homo naledi
Homo neanderthalensis
Homo rhodesiensis
Homo helmei
Homo heidelbergensis
Homo antecessor
Homo cepranensis
Homo erectus
Homo erectus georgicus
Homo ergaster
Homo rudolfensis
Homo habilis
Homo gautengensis
Kenyanthropus
Kenyanthropus platyops
Paranthropus
Paranthropus robustus
Paranthropus boisei
Paranthropus aethiopicus
Australopithecus
Australopithecus sediba
Australopithecus garhi
Australopithecus africanus
Australopithecus bahrelghazali
Australopithecus afarensis
Australopithecus anamensis
Ardipithecus
Ardipithecus ramidus
Ardipithecus kadabba
Orrorin
Orrorin tugenensis
Sahelanthropus
Sahelanthropus tchadensis